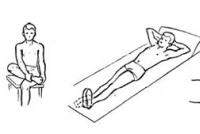
Реакция первичного окончания мышечного веретена. Рефлекс на растяжение мышцы. Рефлекс на растяжение мышцы Как называются рефлексы на растяжение мышцы
III. Примеры двигательных рефлексов.
1. Мышечные рефлексы растяжения и торможения.
Рассмотрим мышечный рефлекс растяжения. Он предназначен для того, чтобы регулировать положение конечностей, обеспечивать неподвижное положение тела, поддерживать тело во время того, как оно стоит, лежит или сидит. Этот рефлекс поддерживает постоянство мышечной длины. Растяжение мышцы вызывает активацию мышечных веретен и сокращение, т. е. укорочение мышцы, противодействующей ее растяжению. Например, когда человек сидит, происходит растяжение мышц брюшного пресса и повышение их тонуса, противодействующее сгибанию спины. И наоборот, слишком сильное сокращение мышцы ослабляет стимуляцию ее рецепторов растяжения, мышечный тонус ослабевает
Рассмотрим прохождение нервного импульса по рефлекторной дуге. Следует сразу отметить, что мышечный рефлекс растяжения относится к простейшим рефлексам. Он проходит непосредственно от сенсорного нейрона к двигательному (рис.1). Сигнал (раздражение) поступает от мышцы на рецептор. По дендритам сенсорного нейрона импульс проходит в спинной мозг и там кратчайшим путем проходит в двигательный нейрон соматической нервной системы, а далее по аксону двигательного нейрона импульс попадает на эффектор (мышцу). Таким образом, осуществляется мышечный рефлекс растяжения.
Рис.1. 1 – мышца; 2 – мышечные рецепторы; 3 – сенсорный нейрон; 4 – двигательный нейрон; 5 – эффектор.
Другим примером двигательного рефлекса является рефлекс торможения. Он возникает как ответ на действие рефлекса растяжения. Тормозная рефлекторная дуга включает два центральных синапса: возбуждающий и тормозной. Можно сказать, что в данном случае мы наблюдаем работу мышц-антагонистов в паре, например, сгибателя и разгибателя в суставе. Мотонейроны одной мышцы тормозятся во время активации другого компонента пары. Рассмотрим сгибание коленного сустава. При этом мы наблюдаем, растяжение мышечных веретен разгибателя, что усиливает возбуждение мотонейронов и торможение мотонейронов сгибателя. Кроме того, уменьшение растяжения мышечных веретен сгибателя ослабляет возбуждение гомонимных мотонейронов и реципрокное торможение мотонейронов разгибателя (растормаживание). Под гомонимными мотонейронами мы понимаем все те нейроны, которые посылают аксоны к одной и той же мышце или возбуждают ту мыщцу, от которой берет начало соответствующий путь от перефирии к нервному центру. А реципрокное торможение – это процесс в нервной симстеме, основанный на том, что по одному и тому же афферентному пути осуществляется возбуждение одних групп клеток и торможение других групп клеток через втавочные нейроны. В конечном счете, мотонейроны разгибателей возбуждаются, а сгибателей – сокращаются. Таким образом, происходит регуляция длины мышцы.
Рассмотрим прохождение нервного импульса по рефлекторной дуге. Нервный импульс зарождается на мышце разгибателя и по аксонам сенсорного нейрона проходит в спинной мозг. Так как данная рефлекторная дуга относится к дисинаптическому типу, то импульс раздваивается, одна часть попадает на мотонейрон разгибателя для поддрежания длины мышцы, а другая – на мотонейрон сгибателя, происходит торможение разгибателя. Затем каждая часть нервного импульса переходит на соответствующий эффектор. Либо, в спинном мозге возможен переход на мотонейрон сгибателей коленного сустава через тормозные синапсы, которые позволяют изменять длину мышцы, а затем по двигательным аксонам выход на концевые пластинки (эффектор, скелетную мышцу). Возможны два других варианта, когда возбуждение воспринимает рецептор сгибателя, тогда рефлекс проходит по такому же пути.

ОРис.2 1. Мышца разгибатель. 2. Мышца сгибатель. 3. Мышечный рецептор. 4. Сенсорные нейроны. 5. Тормозные интернейроны. 6. Двигательный нейрон. 7. Эффектор
Познакомимся теперь с более сложными рефлексами.
2. Сгибательный и перекрестный разгибательный рефлекс.
Как правило, рефлекторные дуги включают в себя два и более последовательно связанных нейронов, т. е. являются полисинаптическими.
Примером может служить защитный рефлекс у человека. При воздействии на конечность, она отдергивается путем сгибания, например, в коленном суставе. Рецепторы данной рефлекторной дуги находятся в коже. Они обеспечивают движение, направленное на удаление конечности от источника раздражения.
При раздражении конечности происходит сгибательный рефлекс, конечность отдергивается, а противоположенная выпрямляется. Так происходит в результате прохождения импульса по рефлекторной дуге. Воздействуем на правую ногу. От рецептора правой ноги по аксонам сенсорного нейрона импульс попадает в спинной мозг, далее он направляется на четыре разных интернейроновых цепи. Две цепи идут на мотонейроны сгибателя и разгибателя правой ноги. Происходит сокращение мышцы сгибателя, а разгибатель расслабляется под воздействием тормозных интернейронов. Мы отдергиваем ногу. В левой ноге происходит расслабление мышцы сгибателя и сокращение мышцы разгибателя под воздействием возбуждающего интернейрона.

РисЧерные – тормозные интернейроны; красные возбуждающие. 2. Двигательные нейроны. 3.Эффекторы расслабленных мышц сгибателя и разгибателя. 4. Эффекторы сокращенных мышц сгибателя и разгибателя.
3. Сухожильный рефлекс.
Сухожильные рефлексы служат для поддержания постоянства напряжения мышцы. У каждой мышцы есть две регулирующие системы: регуляция длины, с помощью мышечных веретен в роли рецепторов и регуляция напряжения, в роли рецепторов в данной регуляции выступают сухожильные органы. Отличие системы регуляции напряжении от системы регуляции длины, в которой задействованы мышца и ее антагонист , заключается в использовании сухожильным рефлексом мышечного тонуса всей конечности.
Развиваемая мышцей сила зависит от её предварительного растяжения, скорости сокращения, утомления. Отклонение от мышечного напряжения от нужной величины регистрируется сухожильными органами и корректируется сухожильным рефлексом.
Рецептор (сухожилие) данного рефлекса находится в сухожилии конечности на конце мышцы сгибателя или мышцы разгибателя. Оттуда, по аксонам сенсорного нейрона сигнал проходит в спинной мозг. Там сигнал может пройти по тормозному интернейрону на двигательный нейрон разгибателя, который отправит сигнал на мышцу разгибатель, для поддержания мышцы в напряжении. Также сигнал может пойти на возбуждающий интернейрон, который отправит сигнал через двигательный аксон на эффектор сгибателя, для изменения напряжения мышцы и совершения определенного действия. В случае, когда возбуждение воспринимает рецептор (сухожилие) сгибателя, сигнал проходит через аксон сенсорного нейрона на интернейрон, а оттуда, на двигательный мотонейрон, который по аксонам двигательного нейрона посылает сигнал в мышцу сгибателя. В рефлекторной дуге сгибателя возможен путь только через тормозной интернейрон.

Рис.Сухожильный рецептор. 2. Сенсорный нейрон. 3. Тормозной интернейрон. 4. Возбуждающий интернейрон. 5. Двигательный нейрон. 6. Рецептор.
В XVII веке математик и философ Рене Декарт (Descartes R.) в своём "Трактате о человеке" попытался объяснить деятельность мозга в понятиях бурно развивавшейся в то время механики. Он предположил существование "животных духов" в форме то ли особого рода жидкости, то ли подвижного пламени, которые циркулируют в организме. Достигнув мозга, эти духи отражаются, подобно лучам света, от полостей желудочков или от занимающей в мозгу центральное положение шишковидной железы. Отражённые духи действуют на моторные пути, а затем и на мышцы, заставляя их сокращаться. Эта наивная модель у наших просвещённых современников может вызвать лишь ироническую улыбку, но её роднит с теперешним пониманием рефлекса представление об отражении, отражённых реакциях (лат. reflection – отражение). Рефлексы и сегодня принято объяснять, как проявление отражательной деятельности центральной нервной системы на различные раздражения.
В 1863 году, т. е. в пору утверждения в России радикального материализма (или нигилизма, выразителем которого был, например, запоминающийся персонаж Тургенева – Базаров), И. М. Сеченов объяснял это так: "Чистые рефлексы, или отражённые движения, всего лучше наблюдать на обезглавленных животных и преимущественно на лягушке, потому что у этого животного спинной мозг, нервы и мышцы живут очень долго после обезглавливания. Отрежьте лягушке голову и бросьте её на стол. В первые секунды она как бы парализована, но не более как через минуту вы видите, что животное оправилось и село в позу, которую оно обыкновенно принимает на суше, т. е. сидит, поджавши под себя задние лапы и опираясь в пол передними. Дотроньтесь до кожи, лягушка шевельнётся и опять покойна. Щипните посильнее, и она, пожалуй, сделает прыжок, как бы стараясь убежать от боли. Механизм этих явлений чрезвычайно прост: от кожи к спинному мозгу тянутся чувствующие нервные нити, а из спинного мозга выходят к мышцам нервы движения; в самом же спинном мозгу обоего рода нервы связываются между собою при посредстве так называемых нервных клеток. Целость всех частей этого механизма совершенно необходима. Перережьте чувствующий или движущий нерв или разрушьте спинной мозг – и движения от раздражения кожи не будет. Этого рода движения называются отражёнными на том основании, что здесь возбуждение чувствующего нерва отражается на движущем."
Из приведённой цитаты следует, что полтора столетия назад были изучены некоторые стереотипные двигательные реакции в ответ на раздражения и уже тогда не вызывала сомнения необходимость соединений между чувствительными и двигательными нервами, хотя и не были ещё открыты синапсы. Из этого же описания следует, что для многих стереотипных реакций даже не требуется головной мозг. Лягушек, лишённых головного мозга, называют спинальными, и все наблюдаемые у них рефлексы являются исключительно спинномозговыми, т. е. замыкаются через спинной мозг. Но приведённая выше цитата взята из работы Сеченова "Рефлексы головного мозга", где он пытался представить любую деятельность больших полушарий мозга, в том числе – психическую, как рефлекторную. Эта гипотеза была умозрительной и никак не подтверждалась экспериментальными данными.
Рефлекс можно определить как закономерную целостную стереотипную реакцию организма на изменения внешней среды или внутреннего состояния, которая осуществляется при обязательном участии центральной нервной системы. Рефлекс обеспечивается объединением афферентных, вставочных и эфферентных нейронов, составляющих рефлекторную дугу.
Можно привести множество примеров стереотипных рефлекторных реакций, которые обнаруживаются у всех людей. Так, например, человек, нечаянно взявший сильно нагретый предмет, немедленно отдёргивает от него руку, а наступивший босой ногой на острый камешек или колючку – тотчас сгибает ногу. И в том, и в другом случае сгибание конечности позволяет избежать ещё большего повреждения, и то, и другое – примеры безусловного защитного рефлекса. Такие рефлексы являются врождёнными и видовыми, поскольку обнаруживаются у всех представителей одного вида. Такими же врождёнными безусловными рефлексами следует считать мигание в ответ на попадание соринки на роговицу глаза и кашель в связи с образованием мокроты в верхних дыхательных путях или проникновением в них инородного тела: и мигание, и кашель способствуют удалению инородных тел, тем самым предупреждая повреждение роговицы или слизистой оболочки дыхательных путей.
Наряду с защитными можно выделить большую группу пищевых безусловных рефлексов, обеспечивающих повышение секреции пищеварительных желёз и усиление моторики желудка и кишечника в ответ на поступление пищи в рот, а затем в желудок и кишечник. К терморегуляционным рефлексам можно отнести расширение сосудов кожи и обильное выделение пота у находящегося в бане человека: таким способом организм пытается предупредить повышение температуры тела. Одышка и учащение пульса у человека, пробежавшего стометровку или быстро поднявшегося на девятый этаж, тоже возникают рефлекторно. При физической работе в организме повышается образование углекислого газа и увеличивается потребление кислорода, а изменённое значение параметров этих газов в крови рефлекторно стимулирует работу сердца и лёгких. Посредством рефлекторной регуляции организм может быстро защищаться от вредных воздействий среды, глотать и переваривать проглоченную пищу, сохранять постоянство параметров внутренней среды и одновременно регулировать их, приспосабливая то к покою, то к различным видам деятельности.
В зависимости от происхождения все рефлексы можно подразделить на врождённые или безусловные и приобретённые или условные. В соответствии с их биологической ролью можно выделить защитные или оборонительные рефлексы, пищевые, половые, ориентировочные и т. д. По локализации рецепторов, воспринимающих действие раздражителя, различают экстероцептивные, интероцептивные и проприоцептивные; по расположению центров – спинномозговые или спинальные, бульбарные (с центральным звеном в продолговатом мозгу), мезенцефальные, диэнцефальные, мозжечковые, корковые. По различным эфферентным звеньям можно различать соматические и вегетативные рефлексы, а по эффекторным изменениям – мигательные, глотательные, кашлевые, рвотные и т. д. В зависимости от характера влияния на деятельность эффектора можно говорить о возбуждающих и тормозных рефлексах. Любой из рефлексов можно классифицировать по нескольким различительным признакам.
Если лапку спинальной лягушки опустить в стакан с раствором кислоты, то она очень скоро, через 2-3 секунды, согнёт её, чтобы вынуть из кислоты, раздражающей чувствительные нервные окончания в коже. По происхождению это безусловный рефлекс, по биологической роли – защитный, по характеру движения – сгибательный, по локализации рецепторов – экстероцептивный (поскольку реагирующие на раздражитель рецепторы находятся в коже, т. е. являются наружными), по уровню замыкания или расположения нервного центра – спинномозговой.
Если сдавить лапку спинальной лягушки пинцетом, то она попытается её вырвать, совершая все необходимые для этого движения, причём их интенсивность окажется пропорциональной силе раздражения: чем сильнее он действует, тем больше нейронов и мышечных волокон возбуждается, тем энергичней ответная реакция на него и наоборот. Сопоставим это обстоятельство с термином рефлекс (от лат. reflexus – отражённый) и обратим внимание на то, что рефлекс – реакция приспособительная, она всегда направлена на восстановление равновесия, нарушенного меняющимися условиями среды. Характер рефлекторного ответа зависит от двух особенностей раздражения: силы стимула и места, на которое он действует.
Спинальная лягушка регулярно сбрасывает со своей кожи кусочки бумаги, смоченные раствором кислоты, причём использует ту лапку, которой удобнее всего стряхнуть бумажку. Таким образом, в её действиях обнаруживается координация, несмотря на отсутствие головного мозга. Следовательно, такая координация предусмотрена уже самим механизмом рефлекса.
Рефлекторные ответы стереотипны: повторное действие одного и того же раздражителя на один и тот же участок тела сопровождается одним и тем же ответом и, если такой ответ обнаруживается у одной лягушки, то и у остальных он оказывается точно таким же. Из этого следует, что рефлексы являются видовыми реакциями. которым не надо обучаться, поскольку они относятся к врождённым способам поведения и вся программа рефлекса записана в генетическом коде каждой особи.
У интактной, т. е. неповреждённой лягушки помимо вышеперечисленных можно обнаружить рефлекс перевёртывания, состоящий в том, что положенное на спину животное довольно быстро возвращается к более естественному для себя положению. Спинальная лягушка переворачиваться не может, что позволяет сделать вывод о расположении центра рефлекса перевёртывания в головном мозгу. Если прикоснуться мягкой бумажкой или кисточкой к роговице глаза лягушки, то она немедленно втянет глаз и закроет веко: центр этого защитного роговичного рефлекса также находится в головном мозгу. В зависимости от того, в каком регионе головного мозга происходит переключение возбуждения с афферентных сенсорных путей на эфферентные, можно различать рефлексы продолговатого, среднего мозга, мозжечка и т. д. При разрушении любого необходимого для воспроизведения рефлекса звена: чувствительного, двигательного или центрального, рефлекторный ответ всегда исчезает.
Рефлексы являются составной частью многих сложных регуляторных процессов: они, например, играют важную роль в произвольных действиях человека. Элементарные дуги спинальных рефлексов посредством проводящих путей взаимодействуют с высшими центрами головного мозга. В соответствии с принципами биокибернетики к классическим компонентам рефлекса (стимул Þ нервный центр Þ ответ) следует добавить обратную связь, т. е. механизм предоставления информации о том, удалось или нет с помощью рефлекторной реакции приспособиться к изменениям среды и насколько эффективным оказалось приспособление:
Рефлекторная дуга или рефлекторный путь представляет собой совокупность образований, необходимых для осуществления рефлекса (Рис. 7.1).

В неё входит цепь соединённых посредством синапсов нейронов, которая передаёт нервные импульсы от возбуждённых стимулом чувствительных окончаний к мышцам или секреторным железам. В рефлекторной дуге выделяют следующие компоненты:
1. Рецепторы – высокоспециализированные образования, способные воспринять энергию раздражителя и трансформировать её в нервные импульсы. Различают первичночувствующие рецепторы, которые представляют собой немиелинизированные окончания дендрита чувствительного нейрона, и вторичночувствующие: специализированные эпителиоидные клетки, контактирующие с сенсорным нейроном. Все рецепторы можно подразделить на внешние или экстерорецепторы (зрительные, слуховые, вкусовые, обонятельные, осязательные) и внутренние или интерорецепторы (рецепторы внутренних органов), среди которых полезно выделить проприоцепторы, находящиеся в мышцах, сухожилиях и суставных сумках. Область, занимаемая рецепторами, которые принадлежат одному афферентному нерву (нейрону) называется рецептивным полем этого нерва (нейрона). Действие порогового раздражителя на рецептивное поле приводит к возникновению специализированного рефлекса.
2. Сенсорные (афферентные, центростремительные) нейроны, проводящие нервные импульсы от своих дендритов в центральную нервную систему. В спинной мозг сенсорные волокна входят в составе задних корешков.
3. Интернейроны (вставочные, контактные) находятся в центральной нервной системе, получают информацию от сенсорных нейронов, перерабатывают её и передают эфферентным нейронам. В спинном мозгу тела вставочных нейронов находятся преимущественно в задних рогах и промежуточной области.
4. Эфферентные (центробежные) нейроны получают информацию от интернейронов (в исключительных случаях от сенсорных нейронов) и передают рабочим органам. Тела эфферентных нейронов расположены в центральной нервной системе, а их аксоны выходят из спинного мозга в составе передних корешков и относятся уже к периферической нервной системе: они направляются либо к мышцам, либо к внешнесекреторным железам. Управляющие скелетными мышцами двигательные нейроны спинного мозга (мотонейроны) находятся в передних рогах, а вегетативные нейроны – в боковых рогах. Для обеспечения соматических рефлексов достаточно одного эфферентного нейрона, а для осуществления вегетативных рефлексов необходимо два: один из них располагается в центральной нервной системе, а тело другого находится в вегетативном ганглии.
5. Рабочие органы или эффекторы представляют собой либо мышцы, либо железы, поэтому рефлекторные ответы в конечном счёте сводятся или к мышечным сокращениям (скелетных мышц, гладких мышц сосудов и внутренних органов, сердечной мышцы), или к выделению секретов желёз (пищеварительных, потовых, бронхиальных, но не желёз внутренней секреции).
Благодаря химическим синапсам возбуждение по рефлекторной дуге распространяется только в одном направлении: от рецепторов – к эффектору. В зависимости от количества синапсов различают полисинаптические рефлекторные дуги, в состав которых входит не менее трёх нейронов (афферентный, интернейрон, эфферентный), и моносинаптические, состоящие лишь из афферентного и эфферентного нейронов. У человека моносинаптические дуги обеспечивают воспроизведение только рефлексов растяжения, регулирующих длину мышц, а все остальные рефлексы осуществляются с помощью полисинаптических рефлекторных дуг.
7.4. Нервные центры
В соответствии с классической традицией представление о нервных центрах рефлексов составляет сердцевину всей рефлекторной теории. Под нервным центром понимают функциональное объединение интернейронов, участвующих в осуществлении рефлекторного акта. Они возбуждаются притоком афферентной информации и адресуют свою выходную активность эфферентным нейронам. Несмотря на то, что нервные центры тех или иных рефлексов находятся в определённых структурах мозга, например в спинном, продолговатом, среднем и т. д., их принято считать функциональными, а не анатомическими объединениями нейронов. Дело в том, что многие интернейроны способны участвовать в замыкании не одной, а нескольких рефлекторных дуг, т. е. они могут поочерёдно входить в состав то одного, то другого центра.
Сформулировавший классические принципы рефлекторной теории Чарлз Шеррингтон (Sherrington C. S.) не был склонен их абсолютизировать, что видно хотя бы из следующей цитаты: "Возможно, "простой рефлекс" является чисто абстрактным понятием, так как все части нервной системы связаны воедино и, вероятно, ни одна из них не в состоянии участвовать в какой-либо реакции, не воздействуя и не испытывая воздействия со стороны других частей, причём вся система, несомненно, никогда не находится в состоянии полного покоя. Однако понятие "простая рефлекторная реакция" оправдано, хотя и несколько проблематично."
Центры спинномозговых двигательных рефлексов испытывают влияние двигательных центров ствола мозга, которые, в свою очередь, подчиняются командам нейронов, входящих в состав ядер мозжечка, подкорковых ядер, а также пирамидным нейронам моторной коры. На каждом иерархическом уровне существуют локальные сети нейронов, по которым возбуждение может циркулировать, таким образом сохраняя информацию в пределах этого уровня. Нейроны разных уровней контактируют друг с другом, оказывая возбуждающее или тормозное действие. За счёт конвергенции и дивергенции в процесс переработки информации вовлекается дополнительное количество нейронов, что повышает надёжность функционирования иерархически организованных центров.
Свойства центров целиком определяются деятельностью центральных синапсов. Именно поэтому возбуждение через центр передаётся только в одном направлении и с синаптической задержкой. В центрах происходит пространственная и последовательная суммация возбуждения, здесь возможно усиление сигналов и трансформация их ритма. Феномен посттетанической потенциации демонстрирует пластичность синапсов, их способность изменять эффективность передачи сигналов.
Шеррингтон изучал эти рефлексы на собаках, мозг которых перерезался на разных уровнях: например, между продолговатым мозгом и спинным или между верхними и нижними буграми четверохолмия. С помощью таких экспериментальных моделей удалось детально изучить многие двигательные рефлексы спинного мозга и обнаружить принцип субординации в отношениях между спинным и головным мозгом.
Известно, что каждое движение требует координированных действий нескольких мышц: например, для того, чтобы взять карандаш в руку потребуется участие около дюжины мышц, из которых одни должны сокращаться, а другие – расслабляться. Совместно действующие мышцы, т. е. сокращающиеся или расслабляющиеся одновременно называются синергистами, в отличие от противодействующих им мышц-антагонистов. При любом двигательном рефлексе сокращения и расслабления синергистов и антагонистов безупречно координированы друг с другом.
По каким правилам взаимодействуют нейроны, управляющие сокращающимися и расслабляющимися мышцами? Рассмотрим самый простой случай – рефлекс растяжения, впервые обнаруженный Шеррингтоном у собак с перерезанным на уровне среднего мозга стволом. У таких животных возникает т.н. децеребрационная ригидность (лат. rigiditas – жёсткость, оцепенение), которая проявляется резким повышении тонуса всех мышц-разгибателей, поэтому лапы максимально разогнуты, а спина и хвост прогибаются дугой. В норме тонус разгибателей и сгибателей уравновешивается двигательными ядрами ствола мозга, а после перерезки ствола красные ядра среднего мозга, поддерживающие тонус сгибателей, отделяются от спинного мозга и на этом фоне наблюдается стимулирующее влияние вестибулярных ядер на разгибатели. При попытке согнуть лапу такой собаки, а значит растянуть находящиеся в тоническом сокращении мышцы-разгибатели, исследователь обнаруживает в ответ рефлекторно возникающее сопротивление и дополнительное сокращение мышц. При этом выявляются два компонента рефлекса: 1) сначала сильный кратковременный фазный – в ответ на изменение длины мышцы, т. е. в самый момент сгибания и 2) слабый продолжительный тонический – когда принудительно согнутой лапе не позволяют разогнуться, удерживая растянутое состояние мышцы, т. е. её новую длину.
Рефлексы растяжения можно обнаружить и у интактных животных, однако они оказываются слабее, чем у децеребрированных, а их стереотипность будет выражена меньше, что обусловлено характером активирующих и тормозных влияний двигательных центров головного мозга. Как впоследствии стало известно, в ответ на растяжение мышцы внешней силой возбуждаются реагирующие только на изменение длины рецепторы мышечных веретён (Рис. 7.2), которые связаны с особым типом мелких интрафузальных (от лат. fusus – веретено) мышечных волокон.

От этих рецепторов возбуждение передаётся по чувствительному нейрону в спинной мозг, где окончание аксона разделяется на несколько ветвей. Одни ветви аксона образуют синапсы с мотонейронами мышц-разгибателей и возбуждают их, что, естественно, приводит к сокращению мышц: здесь моносинаптический рефлекс – его дуга образована лишь двумя нейронами. Одновременно с этим остальные ветви афферентного аксона активируют деятельность тормозных интернейронов спинного мозга, которые тотчас подавляют активность мотонейронов для мышц-антагонистов, т. е. сгибателей. Таким образом, растяжение мышцы вызывает возбуждение мотонейронов мышц-синергистов и реципрокно тормозит мотонейроны мышц-антагонистов (Рис. 7.3).

Силу, с которой мышцы сопротивляются изменению своей длины, можно определить как мышечный тонус. Он позволяет сохранять определённое положение тела или позу. Сила гравитации направлена на растяжение мышц-разгибателей, а их ответное рефлекторное сокращение этому противодействует. Если растяжение разгибателей увеличивается, например, когда на плечи опускается тяжёлый груз, то и сокращение усиливается – мышцы не позволяют себя растягивать и благодаря этому сохраняется поза. При отклонениях тела вперёд, назад или в сторону определённые мышцы растягиваются, а рефлекторное повышение их тонуса сохраняет необходимое положение тела.
По тому же принципу осуществляется рефлекторная регуляция длины у мышц-сгибателей. При любом сгибании руки или ноги поднимается груз, которым может быть и сама рука или нога, но любой груз – это внешняя сила, стремящаяся растянуть мышцы. И здесь можно обнаружить, что ответное сокращение регулируется рефлекторно в зависимости от величины груза. В этом легко убедиться на практике: попробуйте перекреститься, а потом повторите эти же движения с пудовой гирей в руке, как это делали силачи в старом российском цирке.
Сухожильные рефлексы названы так потому, что их можно вызвать, легко ударив неврологическим молоточком по сухожилию более или менее расслабленной мышцы. От удара по сухожилию такая мышца растягивается и тотчас рефлекторно сокращается. Например, в ответ на удар неврологическим молоточком по сухожилию четырёхглавой мышцы бедра (которое легко нащупать под надколенником) расслабленная мышца растягивается, а возникающее в связи с этим возбуждение рецепторов мышечных веретён распространяется по моносинаптической дуге к этой же мышце, что вызывает её сокращение (Рис. 7.4). Моносинаптические сухожильные рефлексы можно получить на любой группе мышц, независимо от того, являются они сгибателями или разгибателями. Все сухожильные рефлексы возникают при растяжении мышцы (а значит. являются рефлексами растяжения) и возбуждении рецепторов мышечных веретён.

Кроме длины в работающих мышцах рефлекторно регулируется ещё один параметр: напряжение. Когда человек начинает поднимать груз, напряжение в мышцах нарастает до такого значение, чтобы этот груз можно было оторвать от пола, но не более: для подъёма 10 кг не надо напрягать мышцы, как для подъёма 20 кг. Пропорционально росту напряжения усиливается импульсация от проприоцепторов сухожилий, которые называются рецепторами Гольджи (См. рис. 7.2). Это немиелинизированные окончания афферентного нейрона, расположенные между коллагеновыми пучками сухожильных волокон. С ростом напряжения в мышце такие волокна натягиваются и сдавливают рецепторы Гольджи. Нарастающие по частоте импульсы проводятся от них по аксону афферентного нейрона в спинной мозг и передаются тормозному интернейрону, который не даёт мотонейрону возбудиться больше, чем это необходимо (Рис. 7.5).

Длина и напряжение мышц взаимообусловлены. Если, например, вытянутая вперёд рука ослабит напряжение мышц, то уменьшится раздражение рецепторов Гольджи, а сила тяжести станет опускать руку. Это приведёт к растяжению мышц, нарастанию возбуждения интрафузальных рецепторов и соответствующей активации мотонейронов. В итоге произойдёт сокращение мышц и рука вернётся в прежнее положение.
Сто человек из ста, нечаянно прикоснувшиеся рукой к сильно нагретому предмету, немедленно согнут её, что уберегает от ещё большего повреждения. Эта стереотипная защитная реакция возникает раньше, чем осознаётся смысл происшедшего, она обеспечивается врождённым рефлекторным механизмом, в котором участвуют болевые чувствительные окончания, сенсорный нейрон, интернейроны спинного мозга и мотонейроны для мышц-сгибателей. По такому же рефлекторному стереотипу человек, наступивший босой ногой на колючку или острый камешек, немедленно её сгибает. Это эволюционно древний рефлекс: ведь даже лишённая головного мозга лягушка сгибает погружённую в кислоту лапку.

После травматических разрывов спинного мозга у человека сохраняются рефлексы регуляции мышечной длины и напряжения, защитные сгибательные рефлексы, но локомоторные рефлексы у людей, в отличие от четвероногих, не обнаруживаются. Перейдя к прямохождению, человек был вынужден некоторые полномочия спинного мозга передать головному. Тем не менее, эволюционно старые программы ходьбы, автоматизм такого рода деятельности сохранились и у него. Например, когда человек идёт, он редко задумывается о чередующихся движениях своих ног, может разговаривать на ходу, а кое-кто ухитряется даже читать. Но, несмотря на это, после травматического разрыва спинного мозга человек становится совершенно беспомощным, поскольку не может совершить ни одного произвольного движения с помощью мышц, которые управляются мотонейронами, расположенными в спинном мозгу каудальнее места повреждения. Он оказывается неспособным координировать мышечный тонус сгибателей и разгибателей и, соответственно, сохранять вертикальную позу и удерживать равновесие, поскольку необходимые для этого нервные центры позно-тонических рефлексов расположены в стволе мозга (См. главу 10).
Под координацией понимают согласованный порядок деятельности нейронов, образующих нервные центры рефлексов. При любом стереотипном движении, даже самом простом, многие мышцы должны согласованно сокращаться и расслабляться. Так, например, человек, наступивший на колючку и рефлекторно подогнувший ногу, нагружает другую, опорную ногу больше обычного, в связи с чем тонус её разгибателей повышается – этот механизм получил название перекрёстного разгибательного рефлекса (Рис. 7.7).

Чтобы сохранить при этих действиях равновесие, придётся изменить положение головы и туловища, а для этого нужно одни мышцы сократить, а другие расслабить. Все эти сокращения и расслабления мышц должны быть не больше, но и не меньше необходимого в каждой конкретной ситуации, все они должны произойти почти в одно и то же время, но, всё-таки не одновременно, а в определённой последовательности.
Деятельностью каждой мышцы управляет далеко не единственный мотонейрон, который способен иннервировать в ней лишь часть мышечных волокон. Вся группа, необходимых для рефлекторного ответа мотонейронов, располагается, как правило в нескольких сегментах спинного мозга. Они могут активироваться при поступлении в спинной мозг возбуждения от разных чувствительных нейронов, одни из которых несут информацию от интрафузальных рецепторов, другие – от рецепторов Гольджи, третьи – от рецепторов, находящихся в коже (в том числе тактильных, болевых, температурных и т. п.).
Растяжение единственной мышцы приводит к возбуждению нескольких сотен чувствительных нейронов, каждый из которых активирует от 100 до 150 мотонейронов. Этот способ взаимодействия нервных клеток, при котором один нейрон многочисленными ветвями аксона действует на большое количество других нейронов, называется дивергенцией. В противоположность этому группа сенсорных нейронов довольно часто направляет окончания своих аксонов к одним и тем же мотонейронам или интернейронам – такая форма взаимодействия называется конвергенцией (Рис. 7.8). Связи клеток внутри нервного центра предопределены генетически, как и связи центров с определёнными сенсорными нейронами и с определёнными эффекторами. Предопределены функциональные роли возбуждающих и тормозных интернейронов, их место в структуре рефлекторных дуг, их медиаторы и постсинаптические рецепторы.

В образовании всех необходимых связей между афферентными и эфферентными нейронами участвуют многочисленные интернейроны – на их долю в мозгу приходится 99, 98% от общего количества нервных клеток. Среди них есть возбуждающие и тормозные нейроны, аксоны которых могут конвергировать к одним и тем же мотонейронам. Многие интернейроны участвуют в соединении одних и тех же мотонейронов с разными сенсорными нейронами, численность которых превышает количество мотонейронов в 5-10 раз. На этом основании Шеррингтон сформулировал, как закономерность, принцип общего конечного пути, подразумевая под ним один и тот же стереотипный двигательный ответ на разные сенсорные стимулы. Например, одинаковый поворот головы возможен при ориентировочных рефлексах в ответ на зрительный, слуховой или температурный раздражители (И. П. Павлов называл подобные реакции рефлексом "что такое?"). Во всех этих случаях используется один и тот же конечный путь – мотонейроны для шейных мышц, тогда как афферентные звенья рефлексов различны.
В связи с этим, при одновременном действии нескольких раздражителей рефлекторная реакция обнаруживается только на один из них, который оказывается наиболее важным в данный момент. В таких случаях активность одного доминирующего центра на время подавляет возбуждение в других центрах. В начале ХХ века петербургский физиолог А. А. Ухтомский сформулировал представление о доминантных очагах возбуждения.
Координация рефлекторной деятельности – это и согласование активности двигательных центров, расположенных в разных регионах мозга. Они объединены проводящими путями и иерархически организованы. В современной литературе, посвящённой физиологии движения, предпочитают говорить не о рефлекторной, а о программной организации центральной нервной системы. Ходьба, например, осуществляется на основе врождённой программы, но любая врождённая программа в течение жизни может изменяться, приобретать характерные индивидуальные приметы, такие, например, как походка моряка или балерины (См. главу 10).
7.10. Вегетативные рефлексы
Кроме скелетных мышц эффекторами рефлекторных реакций могут быть гладкие мышцы внутренних органов, сердечная мышца и железы внешней секреции. Гладкие мышцы есть в стенках кровеносных сосудов, мелких бронхах, пищеварительном тракте; такого типа мышцы изменяют, например, кривизну хрусталика глаза для фокусирования изображения предмета на сетчатке, суживают или расширяют зрачок, в зависимости от условий освещения.
К железам внешней секреции относятся слюнные и потовые, поджелудочная железа и печень, внешнесекреторными являются клетки, выделяющие желудочный и кишечный сок. Объём выделяемого секрета может регулироваться не только нервными, но и гуморальными механизмами, например, с помощью местных гормонов, но в некоторых случаях рефлекторная регуляция является решающим фактором, как, например, при слюноотделении.
Рефлекторная дуга вегетативных рефлексов в своём эфферентном звене содержит два нейрона. Один из них, преганглионарный, расположен в центральной нервной системе, а тело второго, постганглионарного нейрона находится в вегетативном нервном сплетении – ганглии, расположенным вне ЦНС. Почти все внутренние органы иннервируются и симпатическим, и парасимпатическим отделами вегетативной нервной системы, которые обычно оказывают противоположное влияние на эффектор.
Рецепторы афферентных нейронов могут располагаться в самом эффекторе: например, повышение кровяного давления растягивает стенки аорты и этим возбуждает находящиеся там механорецепторы. Поступающие от этих рецепторов в продолговатый мозг сигналы вызывают снижение активности симпатического отдела, что приводит к понижению давления.
В других случаях изменения активности или тонуса вегетативных центров могут вызываться в связи с раздражением каких либо наружных рецепторов, например, находящихся в коже. Так погружение в холодную воду раздражает холодовые рецепторы кожи, что приводит не только к рефлекторному сужению поверхностных кровеносных сосудов, но и к учащению работы сердца, и к небольшому подъёму кровяного давления в связи с повышением тонуса симпатического отдела.
Регуляцию некоторых этапов пищеварения когда-то рассматривали в качестве примера т.н. цепочечных рефлексов. Поступление пищи в желудок рефлекторно повышает его тонус и стимулирует выделение желудочного сока, начинающего расщепление съеденной пищи. При достижении определённой консистенции пищи возникает особый тип сокращений мускулатуры желудка с одновременным расслаблением привратника – мышечного жома между желудком и двенадцатиперстной кишкой. Вследствие этого порция полупереваренной пищи попадает в двенадцатиперстную кишку, что вызывает сокращение привратника и выделение сока поджелудочной железы, а также желчи из желчного пузыря, усиливаются перистальтические движения кишечника. В свете современных представлений эту последовательную координированную деятельность можно представить как реализацию врождённой программы, предусматривающей определённую последовательность активации нейронных популяций или нервных центров.
7.11. Безусловные и условные рефлексы
Приведённые выше примеры рефлексов объединяет то обстоятельство, что они обнаруживаются у всех здоровых людей (или у всех нормальных животных, принадлежащих к одному виду). Это врождённые, видовые приспособительные стереотипные реакции на изменение среды или внутреннего состояния организма. Такие комплексы приспособительных реакций основаны на произошедшем внутриутробно, в процессе формирования мозга, соединении чувствительных нейронов с определёнными интернейронами, эфферентными нейронами и эффекторами. Такие соединения возможны лишь на основе изначально предусмотренного плана, а такой план является важной составной частью генетического кода.
Отбор приспособительных реакций, заносимых в генетический код, происходил на протяжении всей эволюции. Всякий рождающийся организм наделён готовым минимумом приспособительных реакций на все случаи жизни, они обеспечивают возможность движения, пищеварения, регуляции температуры тела, размножения и т. п. И. П. Павлов назвал такие рефлексы безусловными и противопоставил им рефлексы другого рода, приобретаемые каждым организмом самостоятельно на протяжении индивидуальной жизни – условные рефлексы.
В качестве примера такого рефлекса можно привести выделение слюны у взрослой собаки при одном лишь появлении мяса или при его запахе. У щенка такого рефлекса нет, он возникает лишь после того, как вид пищи и её запах несколько раз совпадут с раздражением этой пищей вкусовых рецепторов полости рта. Здесь происходит трансформация поначалу индифферентных, т. е. безразличных раздражителей, которыми являются внешний вид и запах пищи, в условные раздражители, способные вызвать рефлекторное слюноотделение так же, как это раньше делал только безусловный раздражитель – кусок мяса, стимулирующий вкусовые чувствительные окончания
Похожую ситуацию можно вообразить и для человека. Бывает так, что один лишь вид сервированного стола или запах какого-то любимого блюда вызывает у него обильное слюноотделение. Но невозможно представить, что такое может произойти при виде совершенно незнакомого продукта или при ощущении необычного, нетрадиционного гастрономического запаха.
Другой пример образовавшегося условного рефлекса связан с неприятными последствиями какого-либо действия. Так ребёнок, захотевший пощупать впервые увиденное пламя горящей свечи, обжигает себе пальцы и отдёргивает руку, что в дальнейшем несомненно ограничит его исследовательскую активность, зато убережёт от неприятностей.
Условные рефлексы в соответствии с подкрепляющими их безусловными стимулами могут быть классифицированы, например, как пищевые или оборонительные. Их набор у каждого человека индивидуален, всё определяется только его жизненным опытом. Все условные рефлексы формируются на основе безусловных, используют их двигательные или вегетативные центры, их эфферентные нервы и эффекторы: прибавляется только новые формы отношений между определёнными нервными центрами. Предпосылкой для этого служат реально существующие проводящие пути между этими центрами, возможность изменения эффективности синаптической передачи между определёнными нейронными популяциями и т. п. Формирование условных рефлексов, как новых способов приспособления к окружающей среде, демонстрирует пластичность нервной системы, т. е. её умение приспосабливать схемы врождённых программ поведения к самым разным обстоятельствам.
Всякая рефлекторная деятельность не требует участия в ней сознания. Шеррингтон полагал, что сознание и рефлекторная деятельность находятся в реципрокных отношениях, т. е. рефлекторные реакции происходят бессознательно, а сознательная деятельность уже не является рефлекторной. Это однако не исключает возможность сознательного контроля рефлекторной деятельности: например, болевой сгибательный рефлекс можно сознательно подавить волевым усилием.
Резюме
Рефлексы являются элементарными стереотипными приспособительными реакциями организма. Они осуществляются при обязательном участии центральной нервной системы на основе врождённых схем соединения друг с другом чувствительных нейронов, интернейронов, эфферентных нейронов и эффекторов, образующих рефлекторную дугу. В результате рефлекторных реакций организм может быстро приспосабливаться к изменениям внешней среды или внутреннего состояния. Рефлексы являются важной составной частью происходящих в организме регуляторных процессов. Рефлексы спинного мозга находятся под контролем высших центров головного мозга.
Вопросы для самоконтроля
101. Что из перечисленного ниже не является рефлексом?
А. Мигание в ответ на раздражение роговицы инородным телом; Б. Кашель, вызванный попаданием инородного тела в дыхательные пути; В. Образование антител в ответ на поступление в организм чужеродного белка; Г. Выделение слюны во время пережёвывания твёрдой пищи; Д. Одышка, вызванная тяжёлой физической работой.
102. Что из перечисленного ниже не относится к центральной нервной системе?
А. Тела афферентных нейронов; Б. Тела мотонейронов; В. Интернейроны; Г. Вставочные возбуждающие нейроны; Д. Вставочные тормозные нейроны.
103. Какое звено может отсутствовать в рефлекторной дуге?
А. Рецепторы; Б. Интернейроны; В. Сенсорные нейроны; Г. Эфферентные нейроны; Д. Эффекторы.
104. Что из указанного ниже не является эффектором в рефлекторном ответе?
А. Скелетная мышца; Б. Сердечная мышца; В. Гладкая мышца; Г. Слюнная железа; Д. Фолликулы щитовидной железы.
105. Что из указанного ниже является составной частью нервного центра?
А. Рецепторы; Б. Афферентные нейроны; В. Сенсорные нейроны; Г. Интернейроны; Д. Эффекторы.
106. Какое свойство нервного центра обеспечивает возникновение рефлекторного ответа при ритмической стимуляции одного афферентного входа подпороговыми раздражителями?
107. Каким свойством нервного центра можно объяснить возникновение рефлекторного ответа при одновременном действии подпороговых раздражителей на всю поверхность рецептивного поля?
А. Синаптическая задержка; Б. Трансформация ритма; В. Пространственная суммация; Г. Последовательная суммация; Д. Посттетаническая потенциация.
108. Вслед за ритмической стимуляцией афферентного входа в нервный центр рефлекса в течение некоторого времени наблюдается повышенная эффективность синаптической передачи. С каким свойством нервного центра это может быть связано?
А. Синаптическая задержка; Б. Трансформация ритма; В. Пространственная суммация; Г. Последовательная суммация; Д. Посттетаническая потенциация.
109. Мышца рефлекторно сократилась в ответ на растяжение её внешней силой. Что возбудило её мотонейроны?
А. Афферентные нейроны; Б. Интернейроны спинного мозга; В. Нейроны красных ядер; Г. Нейроны вестибулярных ядер; Д. Нейроны ретикулярной формации.
110. Какой элемент рефлекторной дуги не является обязательным для регуляции мышечного напряжения?
А. Рецепторы Гольджи; Б. Афферентный нейрон; В. Возбуждающий интернейрон; Г. Тормозной интернейрон; Д. Эфферентный нейрон.
111. Что из указанного ниже не используется в рефлекторной дуге, обеспечивающей регуляцию напряжения мышц?
А. Сухожильные рецепторы; Б. Рецепторы Гольджи; В. Рецепторы интрафузальных волокон; Г. Тормозные интернейроны; Д. Всё перечисленное является строго обязательным.
112. В ответ на лёгкий удар неврологическим молоточком по сухожилию четырёхглавой мышцы бедра после короткого латентного периода она сокращается и вследствие этого свободно свисавшая голень приподнимается. Раздражением каких рецепторов обусловлен этот рефлекс?
А. Сухожильных рецепторов; Б. Рецепторов Гольджи; В. Осязательных рецепторов кожи; Г. Болевых рецепторов; Д. Интрафузальных рецепторов.
113. Нечаянно дотронувшийся до сильно нагретого предмета человек немедленно отдёргивает от него руку. Где расположен нервный центр этого рефлекса?
А. Спинной мозг; Б. Ствол мозга; В. Средний мозг; Г. Чувствительный ганглий;
Д. Моторная кора.
114. После изоляции спинного мозга у экспериментального животного возникает т.н. спинальный шок, после прекращения которого можно обнаружить восстановление некоторых форм регуляции двигательных функций. Какая двигательная функция не сможет восстановиться?
А. Сухожильные рефлексы; Б. Рефлексы растяжения мышц; В. Сгибательные рефлексы; В. Произвольные движения конечностей; Д. Ритмические рефлексы.
Когда длина мышечного веретена изменяется внезапно, первичное окончание (но не вторичное) стимулируется особенно мощно. Это избыточное возбуждение первичного окончания называют динамическим ответному что означает чрезвычайно активную реакцию первичного окончания на высокую скорость изменения длины веретена. Даже когда длина веретена увеличивается лишь на долю микрометра и это увеличение осуществляется в течение доли секунды, первичный рецептор передает громадное количество дополнительных импульсов по большим сенсорным нервным волокнам диаметром 17 мкм, но только до тех пор, пока длина действительно увеличивается. Как только увеличение длины прекращается, этот дополнительный прирост импульсного разряда возвращается к гораздо более низкому уровню, чем статический разряд, все еще присутствующий в ответе.
Наоборот, при укорочении веретена происходит прямо противоположное изменение сигнала. Таким образом, первичное окончание посылает чрезвычайно сильные, положительные или отрицательные, сигналы к спинному мозгу, информируя его о любом изменении длины мышечного веретена.
Регуляция интенсивности статических и динамических ответов гамма-мотонейронами. Гамма-двигательные нервы к мышечному веретену можно разделить на два типа: гамма-динамические (гамма-d) и гамма-статические (гамма-s). Первые из них возбуждают главным образом интрафузальные волокна с ядерной сумкой, а вторые возбуждают в основном интрафузальные волокна с ядерной цепочкой. Когда гамма-d волокна возбуждают волокна с ядерной сумкой, динамический ответ мышечного веретена становится чрезвычайно усиленным, тогда как статический ответ почти не меняется.
Наоборот, стимуляция гамма-s волокон, возбуждающих мышечные волокна с ядерной цепочкой, усиливает статический ответ, оказывая лишь незначительное влияние на динамический ответ.
Непрерывный разряд мышечных веретен при нормальных условиях. В норме, особенно на фоне некоторой степени возбуждения гамма-эфферентных волокон, в сенсорных нервных волокнах мышечных веретен постоянно возникают импульсы. Растяжение мышечных веретен увеличивает частоту импульсов, тогда как укорочение веретен снижает ее. Таким образом, веретена могут посылать к спинному мозгу положительные сигналы, т.е. увеличенное количество импульсов, свидетельствующее о растяжении мышцы, или отрицательные сигналы, т.е. количество импульсов ниже нормы, что указывает на отсутствие растяжения мышцы.
Рефлекс на растяжение мышцы
Самым простым проявлением функции мышечных веретен является рефлекс на растяжение мышцы. Всякий раз, когда мышца внезапно растягивается, возбуждение веретен вызывает рефлекторное сокращение крупных мышечных волокон растянутой мышцы и тесно связанных с ней мышц-синергистов.
Нервный контур рефлекса на растяжение . На рисунке изображен основной контур рефлекса на растяжение мышечных веретен. Видно, что проприоцептивное нервное волокно типа 1а, исходящее от мышечного веретена, входит в задний корешок спинного мозга. Затем ветвь этого волокна идет непосредственно к переднему рогу серого вещества спинного мозга и синаптически связывается с передними мотонейронами, которые посылают двигательные нервные волокна к той же мышце, от которой исходят волокна мышечных веретен. Таким образом, существует моносинаптический путь, позволяющий рефлекторному сигналу вернуться с кратчайшей задержкой назад к мышце после возбуждения веретена. Большинство волокон типа II от мышечных веретен заканчиваются на множестве вставочных нейронов серого вещества, и их аксоны проводят сигналы к передним мотонейронам с задержкой или выполняют другие функции.
Механизмы возбуждения проприорецепторов
Значение проприорецепторов
Информация в ЦНС о тонусе мышц и положении тела в пространстве поступает от вестибулярного аппарата, глаз и мышечно-суставных рецепторов (собственных рецепторов или проприорецепторов) скелетных мышц, сухожилий, связок, капсул суставов. Сложные двигательные акты координируются с помощью проприорецепторов (механорецепторов) - мышечных веретен, расположенных в скелетных мышцах, и телец Гольджи, расположенных в сухожилиях.
ЦНС получает информацию от проприорецепторов по спиноталамическим и спиномозжечковым проводящим путям глубокой чувствительности об интенсивности и согласованности сокращений отдельных мышц и мышечных групп, изменении движений в суставах при разных нагрузках. Корковая зона проприоцептивного анализатора расположена в прецентральной извилине лобной доли. Анализируя информацию,полученную от проприорецепторов, ЦНС посылает ответные двигательные импульсы мышцам, целесообразно изменяя характер движений. Благодаря проприорецепторам человек и без помощи зрения вполне ориентирован в положении своего тела и его частей в пространстве, осознает направление движения, степень напряжения мышц, необходимую для выполнения движения и поддержания позы.
Мышечные веретена расположены в толще скелетных мышц параллельно мышечным волокнам. Их количество в каждой мышце зависит как от ее размера, так и от выполняемой функции. Одним концом они прикрепляются к мышце, другим – к сухожилию. Возбуждение в них возникает при удлинении мышечных волокон, сухожилий при расслаблении или пассивном растяжении мышц. Мышечные веретена – рецепторы растяжения. В них при растяжении мышцы увеличивается частота нервных импульсов. При изотоническом сокращении мышцы частота импульсов снижается или прекращается. Сухожильные тельца Гольджи, наоборот, растягиваются и возбуждаются при мышечном сокращении. Импульсы от них по афферентным нервным волокнам поступают в ЦНС. Таким образом, мышечные веретена регистрируют изменение длины мышцы, а рецепторы сухожилий – ее напряжение (тонус).
Импульсы от мышечных веретен при растяжении мышцы поступают к мотонейронам спинного мозга, в результате мышца сокращается. Это самый простой пример рефлекторной дуги, в которую входит один синапс (моносинаптической). Самый известный моносинаптический рефлекс – коленный. Эти рефлексы регулируют длину мышцы. Механизм важен для мышц, поддерживающих позу при беге, ходьбе.
При сокращении мышц возбуждаются сухожильные рецепторы с одновременным торможением мотонейронов спинного мозга той же стороны. Ослабление мышечного тонуса активирует мотонейроны. Таким образом, по рефлекторным дугам сухожильных рецепторов поддерживается постоянно высокий мышечный тонус.